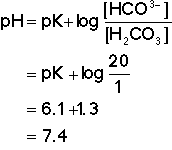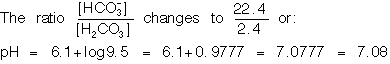There is a normal pH value in each body compartment (i.e. extracellular fluid, plasma, intracellular fluid etc). Intracellular pH is difficult to measure and may vary in different types of cells and in different parts of cells.
pH of the plasma (i.e. pH of the plasma of whole blood = conventional "blood" pH) is controlled at 7.4 (7.35 - 7.45). This section discusses the processes which restore the blood pH to normal if it is displaced.
Changes in plasma pH reflect pH changes in other compartments. When the source of pH change is intracellular the plasma pH change will be in the same direction as the intracellular pH change but of lesser magnitude. When the primary change is in the extracellular fluid the magnitude of any intracellular change will be less than the extracellular change (Van Slyke 1966).
Theoretically, opposite pH changes could occur from shifts of acid or base from one point of the body to another. Proving that such a change has occurred is generally impossible.
K![]() shifts are said to do this but the evidence is nebulous and the
conclusions conflicting. (See Section
8).
shifts are said to do this but the evidence is nebulous and the
conclusions conflicting. (See Section
8).
There are three mechanisms which diminish pH changes in body fluid: buffers; respiratory; renal.
(a) Proteins are the most important buffers in the body.
They are mainly intracellular and include haemoglobin. The plasma
proteins are buffers but the absolute amount is small compared to
intracellular protein. Protein molecules possess basic and acidic
groups which act as H![]() acceptors or donors respectively if H
acceptors or donors respectively if H![]() is added or removed.
is added or removed.
(b) Phosphate buffer
(H2PO4![]() : HP04
: HP04![]() )
is mainly intracellular. The pK of this sytem is 6.8 so that it is
moderately efficient at physiological pH's. The concentration of
phosphate is low in the extracellular fluid but the phosphate buffer
system is an important urinary buffer.
)
is mainly intracellular. The pK of this sytem is 6.8 so that it is
moderately efficient at physiological pH's. The concentration of
phosphate is low in the extracellular fluid but the phosphate buffer
system is an important urinary buffer.
(c)
H2CO2
: HCO3![]() is not an important true buffer system because normal blood pH (7.4)
is so far from its pK (6.1).
H2CO3 and
HCO3
is not an important true buffer system because normal blood pH (7.4)
is so far from its pK (6.1).
H2CO3 and
HCO3![]() are involved in pH control but they are not acting as a buffer system
as defined in Section 2.4. (See also section
3.3.2 and Appendix A3.2).
are involved in pH control but they are not acting as a buffer system
as defined in Section 2.4. (See also section
3.3.2 and Appendix A3.2).
3.2.1. Normal Acid Load. In dealing with the normal acid load from diet and metabolism (carbonic and other acids) the buffers are only involved in diminishing pH changes in the blood as it passes through capillaries (e.g. when CO2 is added in tissue capillaries or removed in pulmonary capillaries). The normal pH of the intracellular and interstitial fluids is maintained not by buffer action but because acids are removed at the same rate as they are added. As there is no change in the quantity of acid in the interstitial fluid or cells with time there is no change in pH. The buffers diminish change in pH due to short term minor physiological disturbances eg. breath-holding (addition of CO2), severe exercise (lactic acid) or during secretion of gastric acid (Rune et al 1968). (See Appendix 3.2.1).
3.2.2. Abnormal Acid Balance. If there is an abnormality in the acid balance, i.e. acid is added faster than it is removed, resulting in a raised level of acid, the change in pH is less than would have occurred if the same imbalance had occurred in a non-buffer solution.
3.2.3. Low Buffer States. In theory a low protein level, (i.e. hypoproteinaemia or anaemia) may make a patient more sensitive to a positive acid balance. This is not recognised clinically. Abnormalities of the buffering system as such do not produce appreciable abnormalities in either the pH status or the acid-base balance because a situation where there were little or no buffers (phosphate and protein) would be incompatible with life.
3.3.1 Normal. In the normal state (at 37°C) when there is no non-respiratory disturbance in pH the carbonic acid level is kept constant in the blood at 1.2meq/l or PaCO2 of 40mmHg (5.3kPa). (PaCO2 x 0.03) = H2CO3 meq/litre.
3.3.2 Effect of Control of
PCO2 in Minimising pH Changes
due to Non-Respiratory Acids or to Bases. Maintenance of the
PaCO2 level (even without compensation, see
3.3.3), is very important in diminishing pH
changes when non-respiratory changes occur. In a closed system where
only true buffering could occur, large changes in
PCO2 would occur when the levels of acids
other than carbonic or bases are changed. For example, if strong acid
were added part of all of the bases in the buffer pairs would be
neutralised. Some HCO3![]() would therefore become
H2CO3 causing the
PCO2 to rise. In the body this rise in
PCO2 would stimulate the respiratory centre
causing a period of hyperventilation which would lower the
PCO2 to normal. If strong base were added some
of the OH
would therefore become
H2CO3 causing the
PCO2 to rise. In the body this rise in
PCO2 would stimulate the respiratory centre
causing a period of hyperventilation which would lower the
PCO2 to normal. If strong base were added some
of the OH![]() would combine with CO2 to give
HCO3
would combine with CO2 to give
HCO3![]() .
.
OH+ CO2
HCO3
The PCO2 would fall. In the body CO2 would be retained to keep the PCO2 from falling.
The control of PCO2 level necessitates either excretion or retention of CO2 by the lungs. This process greatly diminishes the pH change induced by the non-respiratory acids or bases. In effect carbonic acid is being added or taken away to diminish changes that would have been caused by the non-respiratory base or acid respectively. (See Appendix 3.2).
3.3.3 Compensation of Non-Respiratory Disturbances. The respiratory system can also produce rapid compensation for changes in pH by altering the level of PaCO2 . The change in pH alters respiratory control. This causes the alveolar ventilation to alter such that the PaCO2 moves in a direction to cause the pH to return towards normal, i.e. the PaCO2 moves away from normal (40mmHg) in a direction which returns pH towards normal.
In summary, sections 3.3.2 and 3.3.3 state that:
1. By preventing the PaCO2 from changing when other acids or bases are altered, pH changes are much less than would occur if the system were acting as a true buffer system.
2. Respiratory compensation by moving the PaCO2 away from normal in the opposite direction to that which would have occurred in vitro still further reduces pH changes. (See Appendix 3.2).
3.4.1 Normal. The renal system controls the volume and composition of extra-cellular fluid (E.C.F.). It manipulates the E.C.F. electrolytes to maintain the pH at 7.4. In contrast to the rates of change in PCO2 which the respiratory system can produce (minutes) when compensating for pH changes, the renal compensation is slow (days).
3.4.2 Excess Acid or Base (Non-respiratory). If excess acid other than carbonic, or base is added to the internal environemtn, the kidneys excrete them, thus restoring the composition and pH of extracellular fluid to normal. Until the kidney clears the blood of abnormal constituents the pH (assuming the PCO2 is normal) will remain abnormal.
3.4.3 Deficiency of Acid or Base
(Non-respiratory). If a disturbance is due to loss of acid other
than carbonic the kidneys are unable to restore normality unless the
acid deficiency is restored, e.g. in the alkalosis of pyloric
obstruction correction depends on replacement of lost HCl. Correction
of an acidosis due to loss of base (Na![]() or K
or K![]() + HCO3
+ HCO3![]() ), e.g. diarrhoea, requires the administration of Na and/or K salts
from which Na
), e.g. diarrhoea, requires the administration of Na and/or K salts
from which Na![]() or K
or K![]() + HCO3
+ HCO3![]() can be formed.
can be formed.
NOTE: The kidney can correct states of excess but not states of deficiency.
3.4.4 Changes in CO2. If the pH is
low because of a high PaCO2 (acute respiratory
acidosis) the kidneys raise the blood pH towards normal by excreting
acid H![]() + Cl
+ Cl![]() , NH4
, NH4![]() + Cl
+ Cl![]() or 2Na
or 2Na![]() + Cl
+ Cl![]() +
H2PO4
+
H2PO4![]() (see 3.4.5) (Polak
et al 1961). Since the urine has a lower pH than the blood
entering the kidney, the renal venous blood must have a higher
non-respiratory pH than the renal arterial blood. The renal venous
blood then mixes into the systemic circulation and raises the pH of
the systemic blood towards normal. In the body, the
H2CO2 supply is
practically inexhaustible, therefore the following equation moves to
the right as the [H
(see 3.4.5) (Polak
et al 1961). Since the urine has a lower pH than the blood
entering the kidney, the renal venous blood must have a higher
non-respiratory pH than the renal arterial blood. The renal venous
blood then mixes into the systemic circulation and raises the pH of
the systemic blood towards normal. In the body, the
H2CO2 supply is
practically inexhaustible, therefore the following equation moves to
the right as the [H![]() ]
concentration is lowered.
]
concentration is lowered.
H2CO3+ HCO3
Therefore the [HCO3![]() ]
in blood rises (Peters and Van
Slyke, 1931).
]
in blood rises (Peters and Van
Slyke, 1931).
A high pH due to low PaCO2 is probably compensated by renal excretion of base, i.e. NaHCO2 or KHC03.
See Appendix 3.4 for extra discussion of renal control of pH.
3.4.5 Limitation of Acidity of Urine. Limitation of acidity of urine. The kidneys cannot produce a urine pH of much less than 4.4. Strong acids can be removed from the blood and excreted in the urine by:
(a) Reacting with the basic salt of phosphoric acid in the urine without producing a great fall in pH of urine;
H
H
(b) A more slowly developing process is the addition of NH3 (a base) to the urine.
NH3 + H
Therefore pH of the urine falls less when a given quantity of acid is removed from the blood and added to the urine. NH3 is generated from glutamine and amino acids leaving organic acids which are metabolised to CO2 and water, therefore the generation of NH3 does not result in any permanent change in blood pH because the acids produced are easily metabolised and excreted as CO2 by the lung.
3.5.1 Compensation. When a respiratory (PCO2) change occurs in response to a non-respiratory pH disturbance and vice versa, the resulting secondary disturbance in pH is referred to as a compensatory change in response to the primary disturbance.
Compensation of pH disturbances does not completely restore the pH to normal. The pH abnormality which remains after compensation is in the same direction as the primary disturbance unless some complicating factor has intervened.
Some authors refer to the compensatory response as an acidosis or alkalosis whereas others refer only to the primary disturbance as an acidosis or alkalosis. Inserting the terms "primary" or "compensatory" before acidosis or alkalosis should eliminate the communication problem. A primary acidosis (or alkalosis) can be chemically indistinguishable from a compensatory acidosis (or alkalosis). Compensation is not a normal state.
3.5.2 Correction of a acid-base disturbances implies reversal of the chemical cause of the disturbance (e.g. hydrochloric acid, sulphuric acid, lactic acid, CO2). This is necessary before full correction of the disturbance can be said to have occurred.
The mechanism of control of plasma pH at 7.4 is unknown. The receptor which senses changes in pH is unknown, as is the reason why 7.4 is the appropriate pH. Teleologically, the pH at any particular site is that pH which is optimal for enzyme action.
As with most other physiological parameters in the body controlled at particular levels (e.g. haemoglobin level, 14 g%; blood volume, 5 litres), the mechanism of control is unknown. Discussions of how these parameters are controlled, are discussions of how the values are returned to normal if disturbed (e.g. how bone marrow reacts to lowering of haemoglobin level).
The concept of the "normal" pH value is an example of the philosophic problem of induction, i.e. why do all reasonable medical scientists expect and believe that all or almost all stable human organisms will have a blood pH of a particular value?
A classical explanation (Hume) is that it is 'habit or custom' which convinces. This leads to complete scepticism. Bertrand Russell discusses the problem of induction in "The Problems of Philosophy", Chapter 6, 1912 and "The History of Western Philosophy", Chapter 17, Hume, 1946. Karl Popper gives a satisfying and so far not refuted theoretical justification of our expecting further repetition of past events. This he does in among other places in "Objective Knowledge", Chapter 1, 1972.
The concept of normal values and their ranges has been challenged by Schwartz et al, 1973. It is erroneous to assume that if the normal range is the mean ± 2 standard deviations that any value outside this range is importantly abnormal, i.e. that it leads to a decision being made. The probability of a particular value being importantly abnormal varies if the expectation of it being abnormal varies, i.e. pH values of 7.4 or 7.2 have different meaning in say a patient with chronic obstructive airways disease and in a patient in whom no pH abnormality is suspected.
Most texts state that the HCO3![]() : H2CO2 system is an
efficient physiological buffer because the components of the pair are
controlled separately (Pitts,
1974). As it is not a chemical buffer of any reasonable
efficiency at the blood pH use of the term "buffer" in respect to
HCO3
: H2CO2 system is an
efficient physiological buffer because the components of the pair are
controlled separately (Pitts,
1974). As it is not a chemical buffer of any reasonable
efficiency at the blood pH use of the term "buffer" in respect to
HCO3![]() : H2CO2 action
introduces considerable confusion. This is illustrated in the
following example.
: H2CO2 action
introduces considerable confusion. This is illustrated in the
following example.
Plasma has a [HCO3![]() ]
of approximately 24meq/l and
[H2CO2] of
1.2meq/l, hence:
]
of approximately 24meq/l and
[H2CO2] of
1.2meq/l, hence:
If 1.2meq HCl is added to 1 litre of a solution of 24meq
NaHCO2 in water, 1.2meq
HCO3![]() will be converted to
H2CO3, so:
will be converted to
H2CO3, so:
If the H2CO3 is held
constant then 1.2meq HCl causes the
[HCO3![]() ] to diminish by 1.2meq but the
H2CO3 level remains
constant at 1.2meq/l, therefore:
] to diminish by 1.2meq but the
H2CO3 level remains
constant at 1.2meq/l, therefore:
![]()
7.38 is a trivial drop in pH whereas 7.08 is a large change.
Acting as a buffer the HCO3![]() : H2CO2 system would
have allowed a marked change in pH. Chemically the main process which
actually diminished the pH change when HCl was added was the removal
of another weaker acid
(H2CO3) which was
formed when the H
: H2CO2 system would
have allowed a marked change in pH. Chemically the main process which
actually diminished the pH change when HCl was added was the removal
of another weaker acid
(H2CO3) which was
formed when the H![]() of the HCl was neutralised by HCO3
of the HCl was neutralised by HCO3![]() .
.
If base is added to the HCO3![]() : H2CO3 system at pH
7.4 the change in pH is even greater than when an acid is added. If
1.2meq NaOH is added, the [HCO3
: H2CO3 system at pH
7.4 the change in pH is even greater than when an acid is added. If
1.2meq NaOH is added, the [HCO3![]() ]
rises 1.2meq/l or the following equation moves to the right:
]
rises 1.2meq/l or the following equation moves to the right:
H2CO3H
therefore
[H2CO3] falls
to a very low level so that
[HCO3![]() ]
/ [H2CO3] rises
greatly, and therefore pH rises.
]
/ [H2CO3] rises
greatly, and therefore pH rises.
In vivo H2CO3 is
prevented from falling so the ratio
[HCO3![]() ]
/ [H2CO3]
changes only slightly when [HCO3
]
/ [H2CO3]
changes only slightly when [HCO3![]() ]
rises following the addition of Na0H, therefore the pH changes only
slightly. In other words, if a base is added to the blood the main
reason the pH changes very slightly is that a weak acid
(H2CO3) is added to
keep the PaCO2 constant as the
CO2 is used up in partly neutralizing the
added base:
]
rises following the addition of Na0H, therefore the pH changes only
slightly. In other words, if a base is added to the blood the main
reason the pH changes very slightly is that a weak acid
(H2CO3) is added to
keep the PaCO2 constant as the
CO2 is used up in partly neutralizing the
added base:
Na
Inspection of the Siggaard-Andersen nomogram shows greater changes
in pH if the H2CO3
(PCO2) level in blood is altered, than if
similar quantities of strong acid or base are added while the
PCO2 is held at 40mmHg. Altering the level of
H2CO3 by, say 1.2meq in
blood, is not the same as adding 1.2meq
H2CO3. In blood in
vitro raising the level of
H2CO3 1.2meq (80mmHg
PCO2) causes the
[HCO3![]() ]
to rise by 6meq/l (see Appendix
2.4.2 ) therefore in fact 7.2meq
H2CO3 is added to blood
if the PCO2 is raised from 40 to 80mmHg and
the level of H2CO3
raised 1.2meq/l. Adding 7.2meq
H2CO3 to blood in vitro
therefore gives a pH of 7.08, whereas adding 7.2meq HCl when
PaCO2 is 40mmHg gives a pH of 7.29.
]
to rise by 6meq/l (see Appendix
2.4.2 ) therefore in fact 7.2meq
H2CO3 is added to blood
if the PCO2 is raised from 40 to 80mmHg and
the level of H2CO3
raised 1.2meq/l. Adding 7.2meq
H2CO3 to blood in vitro
therefore gives a pH of 7.08, whereas adding 7.2meq HCl when
PaCO2 is 40mmHg gives a pH of 7.29.
Factors other than buffer action which diminish the pH change due to transfer of CO2 as blood passes through capilliaries
Except in the case of CO2, only a small quantity of acid or base is transferred during any one passage of blood through a capillary. When CO2 is transferred two mechanisms other than buffer action reduce pH fluctuations.
a) Reduced haemoglobin anion is a stronger base than oxygenated haemoglobin. Less haemoglobin exists in the ionised form when it is deoxygenated. During the process of deoxygenation CO2 is added. The resulting fall in pH is diminished by the reduced haemoglobin taking up more Hb) Some of the CO2 is carried directly combined with haemoglobin as carbamino-haemoglobin. This is more readily formed by reduced than by oxygenated haemoglobin.
A knowledge of the conventional postulated mechanism of renal
control of pH is assumed in this section (see Relman,
1968 and Pitts, 1974). Text
book descriptions of renal control of pH (Pitts,
1974) are often not consistent with the overall ("black-box")
function of the kidney in transferring acid or base from the blood to
the urine. Briefly, Pitts and others state that pH of blood is
controlled by the components of the
H2CO3 :
HCO3![]() buffer systems being under separate control, i.e. the respiratory
system controls the
[H2CO3] by
controlling PaCO2 and the kidneys control the
HCO3
buffer systems being under separate control, i.e. the respiratory
system controls the
[H2CO3] by
controlling PaCO2 and the kidneys control the
HCO3![]() level by controlling the renal threshold of
HCO3
level by controlling the renal threshold of
HCO3![]() .
The ratio [HCO3
.
The ratio [HCO3![]() ]
: [H2CO3] is,
therefore, controlled and, thus, according to the
Henderson-Hasselbalch equation pH is controlled. As the pH is
controlled so are the ratios of all the other buffer pairs. In this
section I attempt to show [HCO3
]
: [H2CO3] is,
therefore, controlled and, thus, according to the
Henderson-Hasselbalch equation pH is controlled. As the pH is
controlled so are the ratios of all the other buffer pairs. In this
section I attempt to show [HCO3![]() ]
is not a controlled variable but rather a dependent variable. It
appears to me that pH itself is a controlled variable and
[HCO3
]
is not a controlled variable but rather a dependent variable. It
appears to me that pH itself is a controlled variable and
[HCO3![]() ] is dependent on pH and PCO2.
] is dependent on pH and PCO2.
The conventional description of control of non-respiratory
disturbances emphasizes the control of
[HCO3![]() ]
(Pitts, 1974) rather than the
excretion of acid (which must include an anion). It is axiomatic that
if H
]
(Pitts, 1974) rather than the
excretion of acid (which must include an anion). It is axiomatic that
if H![]() is taken from blood and excreted, an anion must also be excreted.
Exchange of H
is taken from blood and excreted, an anion must also be excreted.
Exchange of H![]() for Na
for Na![]() cannot be the full explanation of excretion of H
cannot be the full explanation of excretion of H![]() in the urine. If the blood pH rises by Na
in the urine. If the blood pH rises by Na![]() for H
for H![]() exchange the blood Na
exchange the blood Na![]() + HCO3
+ HCO3![]() level would rise. This Na
level would rise. This Na![]() rise does not occur. Sodium would have to come from an external
source. Excretion of H
rise does not occur. Sodium would have to come from an external
source. Excretion of H![]() = excretion of acid, therefore acid level falls in the blood, and the
level of anion of the particular acid must fall.
= excretion of acid, therefore acid level falls in the blood, and the
level of anion of the particular acid must fall.
The extracellular fluid (E.C.F.) is the source of urinary acid. It
consists of a mixture of equal numbers of anions and cations in
solution. When the E.C.F. becomes more alkaline the total quantity of
basic anion or base (HCO3![]() ,
OH
,
OH![]() etc.) must increase. (This does not apply to the basic anion of a
weak acid if that acid's removal is the cause of the rise in pH, e.g.
in acute hypocapnia [HCO3
etc.) must increase. (This does not apply to the basic anion of a
weak acid if that acid's removal is the cause of the rise in pH, e.g.
in acute hypocapnia [HCO3![]() ]
falls although pH rises). When the pH rises due to loss of acid other
than carbonic, H2CO2
dissociates to H
]
falls although pH rises). When the pH rises due to loss of acid other
than carbonic, H2CO2
dissociates to H![]() and HCO3
and HCO3![]() .
The H
.
The H![]() partially replaces the H
partially replaces the H![]() of the acid which has been removed.
of the acid which has been removed.
The HCO3![]() level in the blood cannot rise without changes in levels of other
anions or cations in the blood. To do so without effects on
Na
level in the blood cannot rise without changes in levels of other
anions or cations in the blood. To do so without effects on
Na![]() ,
Cl
,
Cl![]() or other ions would defy electrical neutrality, i.e. the blood would
become negatively charged and the urine positive. If the
HCO3
or other ions would defy electrical neutrality, i.e. the blood would
become negatively charged and the urine positive. If the
HCO3![]() level in the blood rises either the cation level (Na
level in the blood rises either the cation level (Na![]() )
must rise or the non-basic anion (Cl
)
must rise or the non-basic anion (Cl![]() )
level fall. The Na
)
level fall. The Na![]() level can be raised only by retention of exogenous Na
level can be raised only by retention of exogenous Na![]() if the E.C.F. volume does not fall. The non-basic anion level could
fall by the kidneys excreting Cl
if the E.C.F. volume does not fall. The non-basic anion level could
fall by the kidneys excreting Cl![]() with the H
with the H![]() .
This is the usual way the kidneys extract and excrete acid from the
blood.
.
This is the usual way the kidneys extract and excrete acid from the
blood.
For some years the medical literature stated that Na![]() and Cl
and Cl![]() were not important in the control of blood pH because they are
neither acids nor bases (Smith,
1951). It is now recognised that Cl
were not important in the control of blood pH because they are
neither acids nor bases (Smith,
1951). It is now recognised that Cl![]() is vital in control of pH under some circumstances (i.e. when it is
the anion of hydrochloric acid) (Schwartz
et al, 1968). This should have been obvious from considerations
of electricaly neutrality.
is vital in control of pH under some circumstances (i.e. when it is
the anion of hydrochloric acid) (Schwartz
et al, 1968). This should have been obvious from considerations
of electricaly neutrality.
The kidney cannot increase the
[HCO3![]() ]
in the blood only by reabsorption of
HCO3
]
in the blood only by reabsorption of
HCO3![]() from the glomerular filtrate. Such action would leave the blood level
unchanged if total reabsorption of
HCO3
from the glomerular filtrate. Such action would leave the blood level
unchanged if total reabsorption of
HCO3![]() occurred. HCO3
occurred. HCO3![]() must be generated and [Na
must be generated and [Na![]() ]
or [Cl
]
or [Cl![]() ]
changes must occur as well.
]
changes must occur as well.
The kidney is said to control pH by controlling the renal
threshold of HCO3![]() (Pitts, 1974). In this explanation threshold of
HCO3
(Pitts, 1974). In this explanation threshold of
HCO3![]() rises if PaCO2 rises and vice versa. If this
were the correct explanation the
HCO3
rises if PaCO2 rises and vice versa. If this
were the correct explanation the
HCO3![]() level should fall during the recovery phase of chronic
CO2 retention, because the
HCO3
level should fall during the recovery phase of chronic
CO2 retention, because the
HCO3![]() threshold should now fall. In fact the
[HCO3
threshold should now fall. In fact the
[HCO3![]() ] and the pH remain high until Cl
] and the pH remain high until Cl![]() in some form is given (Polak et
al, 1961).
in some form is given (Polak et
al, 1961).
If a K![]() deficiency is induced in an animal without an accompanying acid-base
disturbance and then K2S04 administered, the pH and
[HCO3
deficiency is induced in an animal without an accompanying acid-base
disturbance and then K2S04 administered, the pH and
[HCO3![]() ] of blood rise (Bleich et al,
1966). This is impossible to explain if
[HCO3
] of blood rise (Bleich et al,
1966). This is impossible to explain if
[HCO3![]() ] is a controlled variable. The explanation is that S04
] is a controlled variable. The explanation is that S04![]() is not retained by the kidney. When it is excreted it must go with a
cation. The kidney has Na
is not retained by the kidney. When it is excreted it must go with a
cation. The kidney has Na![]() , K
, K![]() or H
or H![]() to choose between. As the levels of Na
to choose between. As the levels of Na![]() and K
and K![]() are controlled, the excretion has to be as 2H
are controlled, the excretion has to be as 2H![]() + S04
+ S04![]() or 2(NH4)
or 2(NH4)![]() + S04
+ S04![]() .
As the urine becomes acid the blood must become alkaline and the
[HCO3
.
As the urine becomes acid the blood must become alkaline and the
[HCO3![]() ]
in blood must rise.
]
in blood must rise.
The persisting high [HCO3![]() ]
due to loss of HCl in pyloric stenosis has been explained as a high
threshold for HCO3
]
due to loss of HCl in pyloric stenosis has been explained as a high
threshold for HCO3![]() (Kassirer et al, (a) 1966).
It would occur if HCl (i.e. gastric juice) was removed in a
nephrectomised patient. The blood leaving the stomach would have a
high pH while acid was being secreted. Therefore, the systemic blood
pH would rise, and with it the
[HCO3
(Kassirer et al, (a) 1966).
It would occur if HCl (i.e. gastric juice) was removed in a
nephrectomised patient. The blood leaving the stomach would have a
high pH while acid was being secreted. Therefore, the systemic blood
pH would rise, and with it the
[HCO3![]() ]
(Le Quesne,1961).
Administration of HCl would correct the high
[HCO3
]
(Le Quesne,1961).
Administration of HCl would correct the high
[HCO3![]() ] without intervention by the kidneys (Bradham,
1968). Unless chloride ion in some form is given the pH and
[HCO3
] without intervention by the kidneys (Bradham,
1968). Unless chloride ion in some form is given the pH and
[HCO3![]() ] cannot be corrected (Schwartz
et al, 1968).
] cannot be corrected (Schwartz
et al, 1968).
Conclusion
a) If it can be shown that the kidneys directly control all the electrolytes in the E.C.F. except HCO3b) The kidneys generally can correct only states of excess. Deficiency states have to be corrected by exogenous supplies of raw materials. The kidney is able to correct high levels of Na
Note: Since this section was written (1972) Knud Engel and Paul Kildeberg "Physiological Viewpoint on Clinical Acid-Base Diagnosis", (1977) have written along similar lines. They point out the absence of value in the concepts of bicarbonate reabsorption and renal bicarbonate threshold etc. I think this view is correct. They emphasize the distinction of metabolisable and non-metabolisable acids. I think it is more useful to think about organic and inorganic acids, although all organic acids may not be metabolisable.
The method of describing inputs and outputs which they recommend although consistent with their and my approach is unnecessarily complex in the clinical situation for which the method is advocated.
Schwartz and Cohen (1978) have reviewed several of the paradoxes inherent in the conventional description of renal acidification mechanisms. The approach used in this book is consistent with their hypotheses, i.e. that variation of acid secretion by the kidney occurs in a direction which would appear to be appropriate for pH homeostasis only if this is consistent with other homeostatic mechanisms, i.e. Na + K control. They think, and I agree, that pH control has a low priority in homeostatic mechanisms.
In contrast the conventional approach postulates a H![]() excretion mechanism which is controlled by deviations of blood pH and
that K
excretion mechanism which is controlled by deviations of blood pH and
that K![]() is involved in the H
is involved in the H![]() excretion mechanism.
excretion mechanism.